

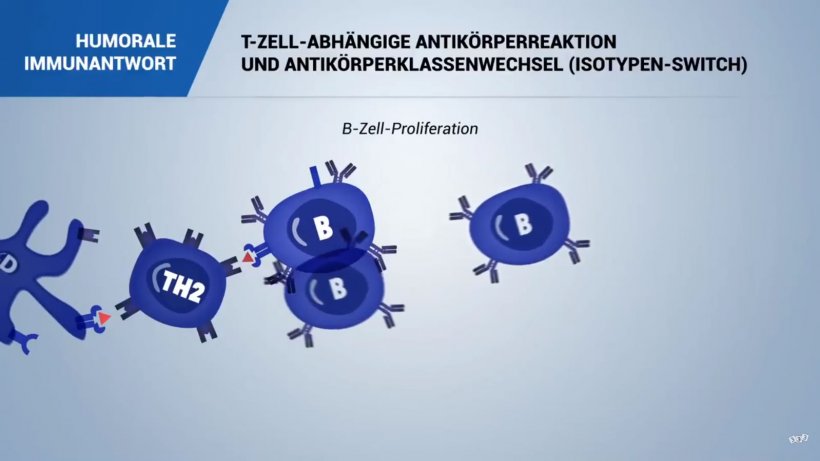
Eine spezifische humorale Immunantwort beginnt, wenn naive B-Lymphozyten ihr spezifisches Antigen in den sekundären Lymphorganen und -geweben erkennen (siehe Pathogenerkennung).
Jeder B-Lymphozyt hat einen Antigenrezeptor (BCR/B-Zell-Rezeptor), ein Oberflächenimmunglobulin (IgM oder IgD), das an spezifische Antigendomänen, auch Determinanten oder Epitope genannt, bindet.

Die Spezifität einer Antigenbindungsstelle hängt von den Genen ab, die für die variablen Domänen (VH und VL) kodieren. Bei Säugetieren kann durch Genrekombination und/oder Genkonversion eine enorme Anzahl an Genen und in der Folge an unterschiedlichen BCR generiert werden, sodass ein Tier in der Lage ist, praktisch alle fremden Antigene zu erkennen.
Nach erfolgter Antigenerkennung wird jeder spezifische B-Lymphozyt aktiviert und es kommt zur klonalen Proliferation und zur Differenzierung in Plasmazellen (oder antikörpersezernierende Zellen/ASC), die in der Lage sind, Immunglobuline (Antikörper) zu synthetisieren und zu sezernieren, und zwar jeweils spezifisch für das entsprechende Epitop.
Die spezifische humorale Immunantwort wird mithilfe der T-Zellen optimiert.
Anfangs sezerniert die B-Zelle IgM in monomerer Form und identischer Antigenspezifität.
Hinsichtlich der Art der Antikörpersekretion werden Antigene als T-Zell-abhängig oder T-Zell-unabhängig klassifiziert, je nachdem, ob für die primäre Immunantwort die Hilfe von T-Zellen notwendig ist oder nicht.
Für bestimmte Antigene können sogenannte T-Zell-unabängige Antigene (z. B. bakterielles Lipopolysaccharid, Flagellin, Pneumokokkenpolysaccharid, etc.) nur IgM-Antikörper mit mäßiger Affinität und keinem B-Zell-Gedächtnis induzieren.
Fast alle fremden Antigene sind T-Zell-abhängige Antigene und benötigen zur vollständigen Aktivierung von B-Lymphozyten ein zusätzliches Signal (Zytokin) von spezifischen T-Helferzellen, die dasselbe Antigen erkennen (T-Zell-B-Zell-Kooperation).
In diesem letzgenannten Fall agiert die B-Zelle als antigenpräsentierende Zelle, die das verarbeitete Antigen den T-Helferzellen präsentiert, und die Antigenerkennung der angeborenen Immunantwort ermöglicht den Antikörperwechsel (Isotyp-Switch) von IgM zu einer anderen Immunglobulinklasse, nämlich IgA, IgE oder IgG, je nach sezerniertem und an der B-Zell-Stimulation beteiligtem TH-Zytokin (IL-4 bzw. IFN-γ). Ein weiteres wichtiges Signal für den Antikörperwechsel ist die Aktivierung des CD40-Rezeptors auf den B-Zellen durch CD154 (CD40L).
Im Rahmen dieser "Hilfe" erfahren reifende B-Zellen eine somatische Hypermutation des Oberflächenimmunglobulins und werden dann mit einem spezifischen Isotyp ausgestattet. Diese B-Lymphozyten werden auf der Basis einer optimalen Ig-Affinität zum Antigen weiter selektiert, indem sie in engen Kontakt mit den follikulären dendritischen Zellen, die das Antigen präsentieren, treten. Nur jene B-Lymphozyten, die eine hohe Antikörperaffinität für das Antigen besitzen und in der Lage sind, dieses zu verarbeiten und zu präsentieren, werden positiv selektiert. Auf diese Art und Weise sind es nur diese B-Lymphozyten, die überleben, proliferieren und zu Plasmazellen differenzieren, die Antikörper einer einzigen Isotyp-Klasse mit einer einzigen Spezifität und hoher Affinität synthetisieren und sezernieren. Dies verbessert die Fähigkeit der Antigenbindung und somit der Neutralisierung und Zerstörung des Pathogens.
Sobald die primäre Immunantwort endet, werden einige dieser bereits durch den ersten Kontakt mit dem Erreger spezialisierten und selektierten B-Zellen und Plasmazellen zu langlebigen Gedächtniszellen, die beim Zweitkontakt mit demselben Antigen (sekundäre Immunantwort) dieselbe Antikörperklasse in höherer Konzentration und mit erhöhter Affinität produzieren.
Zusammenfassend lässt sich sagen, dass bei der primären humoralen Immunantwort zunächst IgM als Einzelklasse in monomerer Form und mit identischer Antigenspezifität sezerniert wird. Danach, nach einigen Tagen, kommt es infolge des Antikörperklassenwechsels (Isotyp-Switch) zur Sekretion einer anderer Antikörperklasse, z.B. IgG. Die sekundäre humorale Immunantwort, die bei einer nachfolgenden Exposition gegenüber demselben Antigen induziert wird, ist hauptsächlich durch die Prooduktion vom IgG-Isotyp, allerdings in höherer Konzentration und mit erhöhter Affinität, gekennzeichnet (Abbildung 1).
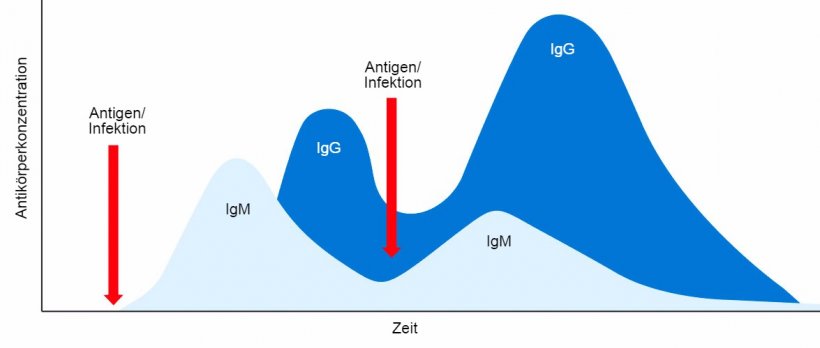
Abbildung 1: Isotyp-Switch sowie primäre und sekundäre Immunantwort bei Zweitkontakt mit demselben Antigen.
Antikörper bekämpfen Erreger mithilfe verschiedener Abwehrmechanismen:
- Hemmung der Infektivität des Pathogens durch direkte Bindung an den Erreger (Neutralisation, Blockierung durch Anhaften, Immun-Ausschluss).
- Aktivierung des alternativen (Opsonisierung) oder klassischen Weges des Komplementsystems mit nachfolgender Lyse des Pathogens.
- Durch Einhüllen des Erregers (oder der infizierten Zelle) versetzen die Antikörper akzessorische Zellen, die das kristallisierbare Fragment (Fc) erkennen, in die Lage, das Pathogen nach dessen Opsonisierung mittels Phagozytose aufzunehmen und abzutöten bzw. infizierte Zellen durch antikörperabhängige zellvermittelte Zytotoxizität (ADCC) abzutöten.
- Regulierung der Immunantworten durch Interaktion mit den Fc-Rezeptoren (FcR).
Fc-Rezeptoren werden von vielen Effektorzellen der angeborenen Immunabwehr exprimiert, z.B. von Mastzellen, Neutrophilen, Monozyten, Makrophagen und dendritischen Zellen. Besonders die Fcγ-Rezeptoren (Fc-Rezeptor für IgG) lösen, wenn sie an Immunkomplexe gebunden sind, die Aktivierung der Zellen des angeborenen Immunsystems und deren Effektorantwort aus (Phagozytose, oxidativer Burst, Zytotoxizität, Freisetzung von vasoaktiven Peptiden und chemotaktischen Attraktantien). FcR auf dendritischen Zellen bewirken das Einfangen des Antigens sowie dessen Verarbeitung und Präsentation an die zytotoxischen T-Zellen und T-Helferzellen.
Umgekehrt sind die auf B-Zellen exprimierten inhibitorischen Fc-Rezeptoren für IgG an der Regulierung der B-Zell-Aktivierung und -Entwicklung sowie am Überleben von Plasmazellen beteiligt.


